
Instigator / Pro
2
1417
rating
27
debates
24.07%
won
Topic
#3480
Macroevolution does not exist.
Status
Finished
The debate is finished. The distribution of the voting points and the winner are presented below.
Winner & statistics
After 1 vote and with 5 points ahead, the winner is...
RationalMadman
Parameters
- Publication date
- Last updated date
- Type
- Standard
- Number of rounds
- 3
- Time for argument
- Two days
- Max argument characters
- 10,000
- Voting period
- One week
- Point system
- Multiple criterions
- Voting system
- Open
Contender / Con
7
1702
rating
573
debates
67.8%
won
Description
Pro:
Has to attack macroevolution
Con:
Has to defend macroevolution
macroevolution:
https://en.wikipedia.org/wiki/Macroevolution
rules:
* Follow sites tos
* do not commit these fallacies https://yourlogicalfallacyis.com/
Round 1
The first problem is that macroevolution takes millions and millions of years to happen. Of course we can't perform any experiment to prove macroevolution because of this. Whatever can be asserted without evidence can be dismissed without evidence. All evolution tested is microevolution, which cannot do many changes. When bacteria evolve in a lab, they are still bacteria, they haven't become a bacteria. This change in bacteria is not enough to prove that bacteria can eventually become humans or other eukaryotes. For macroevolutionary changes to happen, completely new and original information has to be created, which doesn't happen.
The second problem is that many groups of animals appear out of nowhere with no predecessors such as:
Sponges:
- Sponges appear fully formed as the first animals in the deep time scale. An example would be otavia - the "first" true animal
- This animal was not too different from modern sponges.
- Sponges still look the same after a supposed hundreds of millions of years,
Jellyfish and other cnidarians:
- Jellyfish and other cnidarians also appear in the ediacaran fully formed without any predecessors
- These jellyfish have also stayed the same for a supposed millions of years
- Various animals resembling sea pens appear everywhere in the ediacran(they are called Charnia). If these animals are ancestors of modern sea pens, then sea pens haven't actually changed that much for over a supposed hundreds of millions of years
- These animals also appear out of nowhere.
Arthropods:
- Anomalocaris and other anomalocarids
- Trilobites, these creatures appear alongside anomalocarids in the cambrian.
- These animals appear with no predecessor at all
Other invetebrates:
* Scalidophora, this animal includes the modern p*n*s worm(yes it exists) and it also appears right in the beginning(during the cambrian).
Land animals:
- The oldest land animal footprints are a bunch of fully formed tetrapods from Poland and they are older than tiktaalik(an incomplete lobe finned fish fossil).
- These animals are fully formed without predecessors.
Proboscidea:
- proboscideans include mammoths, elephants, mastodons, gomphotheres, and similar
- These animals appear right in the beginning of the paleozoic(during the paleogene) being fully formed.
Dinosaurs:
- Various species of dinosaurs appear in the beginning of the late triassic including eoraptor, coelophyisis, herrerasaurus, plateosaurus, Liliensternus(which looks like a later dilophosaurus), hecodontosaurus, and on and on.
- These animals all appear fully formed without predecessors.
I could go on and on but although these animals don't disprove evolution, they come out of nowhere and disproving macroevolution.
Darwin wrote in his origin of species chapter 6:
Difficulties on the theory of descent with modification. Transitions. Absence or rarity of transitional varieties. Transitions in habits of life. Diversified habits in the same species. Species with habits widely different from those of their allies. Organs of extreme perfection. Means of transition. Cases of difficulty. Natura non facit saltum. Organs of small importance. Organs not in all cases absolutely perfect. The law of Unity of Type and of the Conditions of Existence embraced by the theory of Natural Selection.
A time after the cambrian explosion was discovered, and Darwin had doubts. Even to this day, even after many fossils have been discovered, scientists have yet to solve this doubt.
Pro is implicitly conceding microevolution:
All evolution tested is microevolution, which cannot do many changes. When bacteria evolve in a lab, they are still bacteria, they haven't become a bacteria.
I think that I understand what Pro's angle is and wish to mitigate it to the maximum that I can do in this Round.
Pro's idea of macroevolution being real is that for it to occur, the 'original lifeform' has to have macroevolved from something else. This explains his entire Round 1 logic.
The concept Pro has is faulty and it is actually true that creationism can coincide with macroevolution but not the Biblical Intelligent Design model.
The creationism that can coincide is that we agree the original lifeforms were just mutating cells and that those cells themselves didn't evolve from other things.
Let's now understand what macroevolution does state as a theory and is.
Macroevolution generally refers to evolution above the species level. So instead of focusing on an individual beetle species, a macroevolutionary lens might require that we zoom out on the tree of life, to assess the diversity of the entire beetle clade and its position on the tree.
Macroevolution refers to evolution of groups larger than an individual
Macroevolution encompasses the grandest trends and transformations in evolution, such as the origin of mammals and the radiation of flowering plants. Macroevolutionary patterns are generally what we see when we look at the large-scale history of life.
It is not necessarily easy to “see” macroevolutionary history; there are no firsthand accounts to be read. Instead, we reconstruct the history of life using all available evidence: geology, fossils, and living organisms.
Once we’ve figured out what evolutionary events have taken place, we try to figure out how they happened. Just as in microevolution, basic evolutionary mechanisms like mutation, migration, genetic drift, and natural selection are at work and can help explain many large-scale patterns in the history of life.
The basic evolutionary mechanisms — mutation, migration, genetic drift, and natural selection — can produce major evolutionary change if given enough time.
A process like mutation might seem too small-scale to influence a pattern as amazing as the beetle radiation, or as large as the difference between dogs and pine trees, but it’s not. Life on Earth has been accumulating mutations and passing them through the filter of natural selection for 3.8 billion years — more than enough time for evolutionary processes to produce its grand history.
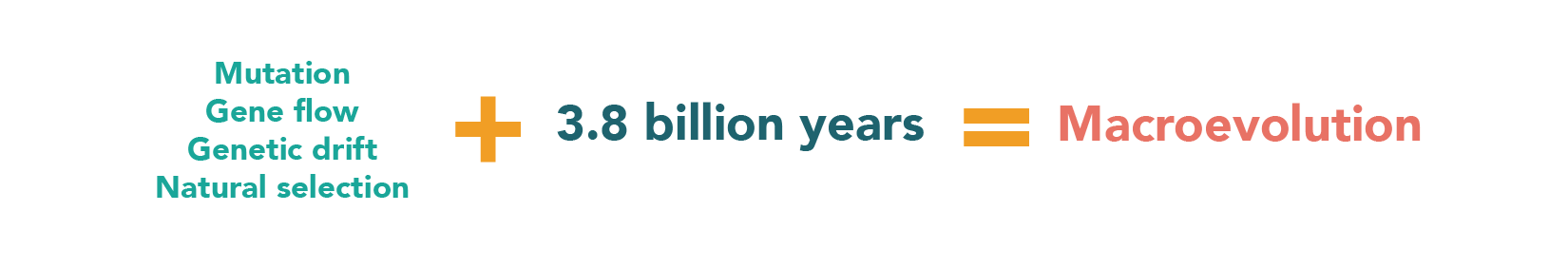{kind=link}
Even if we agree entirely with Pro (and I do agree) that the original lifeform is mysterious in origin and has no macroevolutionary explanation behind it, those original cells and formations which evolved into other species of plant, animal and microorganisms such as bacteria, did so via macroevolution.
What exactly is Pro's explanation for this:
One species becomes two.There are also many examples where new species arise without any barrier—where one population splits in two even though all of its members share the same space. This is called sympatric speciation (from the Greek for “same fatherland”). It was proposed more than a century ago, and has been controversial for much of that time. But scientists have found more and more examples that support the concept, where new wasps, flies, fish and trees evolve side by side. Just last week, news broke about a deadbeat ant that branched off from its parent species, while living in the same colony.The latest intriguing example comes from James Van Leuven and John McCutcheon at the University of Montana. It involves a bacterium called Hodgkinia that split into two distinct species, while living in the cells of an insect. There is no barrier. Sardines in a can have nothing on Hodgkinia. These bacteria are crammed into the same tightly packed microscopic structures, but somehow, they’ve managed to become two distinct species.The two daughter species are like two halves of their ancestor. They’ve each lost genes that the original Hodgkinia had, but they’ve jettisoned different genes. Each compensates for the losses of its sister species. They complement each other perfectly—put them together, and you’d (almost) the complete genome of the ancestor.Van Leuven and McCutcheon made their discovery by studying cicadas—insects known for their ear-splitting songs. About five years ago, they showed that one species of cicada has two bacteria living inside its cells—Sulcia and Hodgkinia. This is pretty normal. Many insects have helpful internal bacteria or “endosymbionts”. In sap-sucking groups like cicadas, these microbes act like dietary supplements, making nutrients that are missing from their diet.Things got strange when Van Leuven and McCutcheon analysed DNA from a South American cicada called Tettigades undata. They found many fragments of Hodgkinia DNA but, try as they might, they couldn’t unite those pieces into a single genome. They always assembled into two separate ones. For simplicity, I’m going to call these H1 and H2.The two genomes belong to different bacteria; they’re never found in the same cell The team confirmed this by using fluorescent molecules designed to label each genome—a yellow one for H1 and a blue one for H2. You can see the results in the image below. Each little dot is a separate bacterium, and each contains either H1 or H2, but never both. (The green dots are Sulcia, and the magenta ones belong to the cicada itself.)Two Hodgkinia species (blue and yellow) in a cicada. Credit: Van Leuven et al, 2014.These two bacteria diverged from a common ancestor, which I’ll call H0, around 5 million years ago. The duo got a good idea of what H0 looked like by studying a closely related species of cicada, which only has one Hodgkinia genome with 137 genes. Out of these, 20 are there in H1 but not H2, and 44 are in H2 but not H1. All of them (except one) are found in one or both of the daughter species.This pattern looks a lot like what happens when a species duplicates its entire genome, as has happened many times in the evolution of flowers, fish, and more. Suddenly, the species carries two copies of each gene. Since it only needs one, the second is free to pick up mutations that disable and destroy it, which is often what happens. The result is a genome with almost the same number of genes, but packed into twice as much material.That’s exactly what Van Leuven and McCutcheon saw in their cicadas. The original Hodgkinia doubled up into two distinct genomes that add back up to the original. But in cases of whole-genome duplication, the doubled-up DNA is still part of the same genome. Not so here; in this case, H1 and H2 are separate entities. The process that created them is a bit like splitting a coin along its edge, so you get a heads-only coin and a tails-only one. There’s no novelty. H1 and H2 don’t do anything that H0 couldn’t already do.That’s abundantly clear if you look at their genes. H0 makes nutrients like vitamin B12 and methionine to feed its cicada host. It devotes many genes to the task, one for each step in the chain of chemical reactions that eventually produce the nutrients. Between them, H1 and H2 can do the same, but neither of them has the complete set of genes for any chain. Neither one alone can give its cicada the nutrients it needs. They have to work together, passing chemicals between them like a production line that snakes between two adjacent factories.How did this complicated set-up evolve? How did H1 and H2 arise from a population of H0 cells that were all living next to one another?It’s hard to say for sure, but McCutcheon has an idea, illustrated in the diagram below. Each Hodgkinia bacterium contains thousands of copies of its genome (the green circles) in the same cell (the black outlines). At some point, one bacterium gets a mutation in just one of its genomes, which breaks one gene (marked in yellow in B). A second bacterium gets a mutation, again in just one genome, which breaks a different gene (marked in blue).
If cross-species evolution doesn't occur and doesn't exist, how is it that the DNA of certain insects prove that deviation occurs and that a species can have dormant DNA from prior species?
Spiders probably evolved about 400 million years ago from thick-waisted arachnid ancestors that were not long emerged from life in water. The first definite spiders, thin-waisted arachnids with abdominal segmentation and silk producing spinnerets, are known from fossils like Attercopus fimbriungus. This spider lived 380 million years ago during the Devonian Period, more than 150 million years before the dinosaurs.
Most of the early segmented fossil spiders belonged to the Mesothelae, a group of primitive spiders with the spinnerets placed underneath the middle of the abdomen (rather than at the end as in 'modern' spiders). They were probably ground dwelling predators, living in the giant clubmoss and fern forests of the mid-late Palaeozoic, where they were presumably predators of other primitive arthropods (like cockroaches, giant silverfish, slaters and millipedes). Silk may have been used simply as a protective covering for the eggs, a lining for a retreat hole, and later perhaps for simple ground sheet web and trapdoor construction.
As plant and insect life diversified so also did the spider's use of silk. Spiders with spinnerets at the end of the abdomen (Opisthothelae) appeared more than 250 million years ago, presumably promoting the development of more elaborate sheet and maze webs for prey capture both on ground and foliage, as well as the development of the safety dragline.
Round 2
Pro is implicitly conceding microevolution:
it's macroevolution, not microevolution.
Macroevolution generally refers to evolution above the species level. So instead of focusing on an individual beetle species, a macroevolutionary lens might require that we zoom out on the tree of life, to assess the diversity of the entire beetle clade and its position on the tree.
This is microevolution. Macroevolution is above the species level - meaning genus, family, order, class, phylum, kingdom, domain, etc. You can't really create a new family in a lab. In other words. You can't empirically prove this definition either
Macroevolution refers to evolution of groups larger than an individual
you are contradicting yourself.
Macroevolutionary patterns are generally what we see when we look at the large-scale history of life.
This is something nobody has ever had a firsthand account of. To test this in a lab will require way too much time to even be possible.
The latest intriguing example comes from James Van Leuven and John McCutcheon at the University of Montana. It involves a bacterium called Hodgkinia that split into two distinct species, while living in the cells of an insect. There is no barrier. Sardines in a can have nothing on Hodgkinia. These bacteria are crammed into the same tightly packed microscopic structures, but somehow, they’ve managed to become two distinct species.
This is microevolution. These bacteria may be 2 new species of bacteria - but they are still bacteria, not amoebas or any new kinds(families).
Spiders probably evolved about 400 million years ago from thick-waisted arachnid ancestors that were not long emerged from life in water. The first definite spiders, thin-waisted arachnids with abdominal segmentation and silk producing spinnerets, are known from fossils like Attercopus fimbriungus. This spider lived 380 million years ago during the Devonian Period, more than 150 million years before the dinosaurs.
This would be proof for intelligent design because blind processes cannot predict the future - and to make a design to last the test time you have to predict the future. For the design of a building to last ages and endure wear and tear, an architect must plan and make a good design. Wouldn't an intelligent being have to do same as the architect?
Key:
MaE = macroevolution
MiE = microevolution
Pro has several misconceptions but let me agree wholeheartedly that MaE is not identical to MiE, microevolution is within a species and slightly the edge of making a breed that becomes somewhat a new species, macroevolution is the part beyond that.
I literally quoted a full explanation of MaE and Pro denies that I defined it.
MaE is the moment a species evolves into a new one. That is MaE, of course Pro is correct that the evolution and emergence of reptiles, mammals etc also is MaE but that is like the severe end of macroevolution. Primates diverging into humans, chimpanzees, orangutans etc is also MaE, not just mammal level meaning species level is MaE but within species the breeds are MiE.
There is nothing about MaE existing that would inherently require it to be able to occur inside of a laboratory. What Pro is saying, however, is untrue as it has happened at the microbiable and insect level within labs.
First, you need to realise that MaE is just MiE on a larger scale the key difference being that while MiE is obvious and conceded by Pro, MaE involves so many changes that the actual species changes (meaning that they've changed so much the the original beings have either maintained or become a species that cannot reproduce fertile offspring with the other, newer one).
The idea of evolution itself with varying degrees of adaptations over time has been proven in a lab:
When it comes to evolution, insects are quick-change artists. In fact, when insects are grown in laboratory conditions, some undergo noticeable adaptations in just a few generations, according to a new review of research published in the Journal of Economic Entomology.Leslie Mertz, Ph.D.“In some cases, we saw shifts in traits of up to 10 percent per generation, so some changes were really moving fast,” says Ary Hoffmann, Ph.D., professor in the University of Melbourne’s Bio21 Molecular Science & Biotechnology Institute. Hoffmann and graduate student Perran Ross co-authored the research, which involved an extensive search of more than 50 scientific studies to examinethe speeds at which laboratory-reared insects adapt, which traits were more often affected, and whether those adaptations improved fitness or weakened it.Overall, the researchers found that constant lab conditions spurred adaptations. “We saw changes in life history, including in how many eggs are produced and how quickly they develop. You might have a strain that produces very few eggs in the wild, but then, over just a few generations in the lab, it’s producing a considerably larger number of eggs,” Hoffmann says. At 10 percent more eggs per generation, numbers can add up fast.This does not mean that every species experiences these changes when grown in a laboratory. Size of the lab population, conditions of the lab environment (e.g., the degree of crowding, temperature), the type of insects being reared, and how long the population has been reared—these can all affect evolutionary rates. “Some traits change quickly; some do not. There’s a lot of variability,” he says.General findings relating to laboratory-reared insects include:
- Mosquitoes and other flies experience adaptations to lab conditions that boost their fitness (e.g., slightly larger size, faster development, more mating). Butterflies and moths, on the other hand, “can go downhill pretty rapidly,” Hoffmann says.
- Morphology doesn’t change much. Although some species may get a little bigger or smaller when raised in a controlled environment, most morphological changes were minor.
- The longer insects are reared in a lab, the lower their tolerance for temperature swings (a trait known as thermal stress resistance). “That’s important because, if a researcher is testing a population to work out whether it is susceptible to extreme temperatures and they are using laboratory stocks rather than wild insects, they could decide it’s not particularly tough when it actually is,” he explains.
- Mating behaviors can adapt quickly. While behavioral adaptations often spur reproduction in the lab, such changes can make lab-grown insects less able to find mates in the field. This can be especially problematic for sterile-release programs.
Understanding the extent of adaptions is important for several reasons, according to Hoffmann. “We rear quite a lot of insects these days for the purpose of biocontrol,” he says, explaining that these include parasitoid wasps that are released to attack certain farm pests. Such biocontrol efforts help reduce reliance on pesticides. “There’s a large number of these lab-reared insects going out in the field, particularly in situations like glass houses [greenhouses], so you want to make sure that they have very high fitness and are going to perform very well. If, however, you are using insects that have started to become adapted to laboratory conditions, then you may find that that fitness and performance decrease, and that can be a real issue.”Likewise, control programs also raise large numbers of sterile male insects, especially mosquitoes, as a way to control the breeding of potentially disease-carrying females. “Millions and millions of sterile males are being released over thousands and thousands of hectares, and those sterile males have to compete with males out in the field. If the lab-reared, sterile males start losing that competitive ability, then you have to start releasing a hell of a lot more of them to have an impact, so it’s a pretty critical issue,” Hoffman says.Those rearing insects would do well to keep in mind the potential for lab adaptations, and they might consider either replenishing their stock at appropriate intervals or cross-breeding the lab population with wild individuals to ensure they have the healthiest and most capable insect stock for release programs and for research studies, Hoffmann advises. “This is extremely important, and just something you really have to do.”In addition to this review study, Hoffmann’s research group is preparing to publish another paper on the rate of adaptation of the mosquito species Aedes aegypti, which spreads yellow fever, Zika, and a variety of other pathogens. “We were expecting a fairly high rate, but it’s at the lower end,” he says. He postulates that the low rates may stem from the fact that these mosquitoes normally live near human habitation, so they are already accustomed to a somewhat artificial environment. “It’s quite an intriguing finding,” he adds.Hoffman and his group are also interested in a couple of parasitoid wasps to be released as a tactic to combat certain leaf-mining insects, and he affirmed that they will indeed be taking adaptation rates into account. He remarks, “Our newly published review paper reinforces the fact that we, like everyone else, have to take lab adaptation seriously and consider it very carefully. So, yes, adaptation rates are very much on our agenda.”
The proofs of MaE exist but rarely have been done inside of a lab as forming a new species and pushing towards MaE level evolution (meaning Pro will say that a new species is still not enough proof).
The formation of new species and genome evolution in stick insects can be repeatable and predictable, researchers from the University of Sheffield have found.As populations diverge to form news species their genomes also diverge, but whether these processes can be repeated and predicted has remained debated.Timema cristinae. Credit: M. Muschick/University of SheffieldA team of scientists from leading universities across the world studied Timema cristinae, a wingless, herbivorous stick insect endemic to California that has repeatedly evolved ecotypes adapted to different host plant species and are in the process of evolving into two unique species.They examined whole-genome divergence between replicate pairs of stick insect populations that have adapted to different host plants and conducted a field experiment to test if repeatable genome evolution was caused by Darwinian natural selection.Their study, the first of its kind, showed the repeated component of the genome evolution occurred by natural selection, while its collective findings demonstrate a repeated element to evolution, even at the genome-wide level and during the complex process of the formation of new species.The study, published in the journal Science today, advances understanding of biological diversification.Dr Patrik Nosil, of the University of Sheffield’s Department of Animal and Plant Sciences, is the senior author of the paper.He said: “As organisms colonise new environments, they either adapt to these environments or go extinct.“Our laboratory studies this process of evolutionary adaptation and its consequences. For example, we test how rapidly adaptation can occur, how repeatable the process is, and what types of genetic changes are involved. In turn, we examine whether adaptation creates new species.“We resequenced the genome of each individual that we collected and looked at which genes were differentiated between populations adapted to different host plants. Because we also conducted an experiment in the field measuring evolution in real time, we gained information on how natural selection is pulling these populations apart."Scott Egan, from Rice University in the USA, said it was previously impossible to conduct this kind of study because of the expense of genomic tests.He added: "The world of genomics is beginning to open up for people like me who don't study model organisms"This is allowing us to address, in new ways, questions that Charles Darwin posed over 150 years ago."
However, what Pro really is wrongly insisting is that I didn't define MaE so let's give a concise one:
evolution that results in relatively large and complex changes (as in species formation)
Round 3
There is nothing about MaE existing that would inherently require it to be able to occur inside of a laboratory. What Pro is saying, however, is untrue as it has happened at the microbiable and insect level within labs.
Need examples of macroevolution occurring in labs.
First, you need to realise that MaE is just MiE on a larger scale the key difference being that while MiE is obvious and conceded by Pro, MaE involves so many changes that the actual species changes (meaning that they've changed so much the the original beings have either maintained or become a species that cannot reproduce fertile offspring with the other, newer one).
Yes but there is no proof that evolution can happen at this large scale. Animals and complex and there are only a few types of base DNA mutation:
- deletion
- mutation
- duplication
- inversion
- insertion
- translocation
There is no mutation that makes complete original information.
Bacteria reproduce very rapidly(every 4 to 20 minutes). The Richard Lenski experiment has only been able to produce the loss of flagella after many years. This is about 3944615.75 generations of bacteria. If you multiply human generations(20 years), you get 78,892,315 years - more than the entire history of hominids, for a few major mutations.
The idea of evolution itself with varying degrees of adaptations over time has been proven in a lab:When it comes to evolution, insects are quick-change artists. In fact, when insects are grown in laboratory conditions, some undergo noticeable adaptations in just a few generations, according to a new review of research published in the Journal of Economic Entomology.Leslie Mertz, Ph.D.
They are still insects. This is microevolution because they are still noticably insects. You haven't specified if they are new families of insects, so I will assume species.
“In some cases, we saw shifts in traits of up to 10 percent per generation, so some changes were really moving fast,” says Ary Hoffmann, Ph.D., professor in the University of Melbourne’s Bio21 Molecular Science & Biotechnology Institute. Hoffmann and graduate student Perran Ross co-authored the research, which involved an extensive search of more than 50 scientific studies to examinethe speeds at which laboratory-reared insects adapt, which traits were more often affected, and whether those adaptations improved fitness or weakened it.Overall, the researchers found that constant lab conditions spurred adaptations. “We saw changes in life history, including in how many eggs are produced and how quickly they develop. You might have a strain that produces very few eggs in the wild, but then, over just a few generations in the lab, it’s producing a considerably larger number of eggs,” Hoffmann says. At 10 percent more eggs per generation, numbers can add up fast.This does not mean that every species experiences these changes when grown in a laboratory. Size of the lab population, conditions of the lab environment (e.g., the degree of crowding, temperature), the type of insects being reared, and how long the population has been reared—these can all affect evolutionary rates. “Some traits change quickly; some do not. There’s a lot of variability,” he says.
Any big percentage of DNA doesn't indicate a new kind. A few small gene changes can make huge difference (as in dog breeds). Other times lots of gene changes can make no difference(as in Algerian mice and house mice).
General findings relating to laboratory-reared insects include:Mosquitoes and other flies experience adaptations to lab conditions that boost their fitness (e.g., slightly larger size, faster development, more mating). Butterflies and moths, on the other hand, “can go downhill pretty rapidly,” Hoffmann says.
These mosquitoes and flies are just super mosquitoes and flies, not new different families or genera. Butterflies/moths that can go downhill rapidly are just butterflies/moths that go down rapidly.
The proofs of MaE exist but rarely have been done inside of a lab as forming a new species and pushing towards MaE level evolution (meaning Pro will say that a new species is still not enough proof).
Because that is not what macroevolution is. A new species in the same family is microevolution, a new species of a new family/genus or above is macroevolution.
In the short description of this debate (available from main 'Debates' Lobby you said ID vs macroevolution.
Where is the ID? You dont even know macroevooution is also speciation
stop trying to sound smart, it doesn't really work.
I said fact, not theory. It's not a theory that a human fetus is a live human being, it's a fact.
A scientific theory is a well-substantiated explanation of some aspect of the natural world, based on a body of facts that have been repeatedly confirmed through observation and experiment. Such fact-supported theories are not "guesses" but reliable accounts of the real world.
I mean it's a scientific fact that a fetus is a human being too but scientists pick and choose to suit agendas, so I don't really think 'fact' vs 'theory' is up to a scientific author to draw.
This is an excellent scientific paper on the subject.
Science and Evolution
Claudia A.M. Russo 1 and Thiago André 2
Abstract
Evolution is both a fact and a theory. Evolution is widely observable in laboratory and natural populations as they change over time. The fact that we need annual flu vaccines is one example of observable evolution. At the same time, evolutionary theory explains more than observations, as the succession on the fossil record. Hence, evolution is also the scientific theory that embodies biology, including all organisms and their characteristics. In this paper, we emphasize why evolution is the most important theory in biology. Evolution explains every biological detail, similar to how history explains many aspects of a current political situation. Only evolution explains the patterns observed in the fossil record. Examples include the succession in the fossil record; we cannot find the easily fossilized mammals before 300 million years ago; after the extinction of the dinosaurs, the fossil record indicates that mammals and birds radiated throughout the planet. Additionally, the fact that we are able to construct fairly consistent phylogenetic trees using distinct genetic markers in the genome is only explained by evolutionary theory. Finally, we show that the processes that drive evolution, both on short and long time scales, are observable facts.
Thank you
Will do when it’s finished.
please vote on this debate if you wish to.
You: "+ Fossil Findings and Noticeable Differences between Alike Species"
An intelligent designer can reuse the same structure to not have to make a new one just like a video game programmer reuses the same code or engine when making a new game.
Not centuries modern darwinist biology appeared in the late 1800s which is pretty recent. I do not deny natural selection or microevolution. Just macroevolution, abiogenesis, and panspermia.
+ Fossil Findings and Noticeable Differences between Alike Species
The thing is at the insect and spider level it is nearly irrefutable.
Wait so you're denying centuries of biological evidence? Interesting...